Revista de la Facultad de Ciencias
Agrarias. Universidad Nacional de Cuyo. Tomo 55(2). ISSN (en línea) 1853-8665.
Año 2023.
Original article
Fragmented
areas due to agricultural activity: native vegetation dynamics at crop
interface (Montecaseros, Mendoza, Argentina)
Áreas
fragmentadas por la actividad agrícola: dinámica de la vegetación nativa en su
interfase con cultivos (Montecaseros, Mendoza, Argentina)
Elena María
Abraham2
1Universidad
Nacional de Cuyo. Facultad de Ciencias Agrarias. Almirante Brown 500. Chacras
de Coria. Luján de Cuyo. Mendoza. Argentina (CPA M5528AHB).
2Instituto
Argentino de Investigaciones de Zonas Áridas (IADIZA). CONICET. Av. Ruiz Leal
s/n. Parque General San Martín. Mendoza. Argentina. (CP 5500).
*avignoni@fca.uncu.edu.ar
Abstract
Plant
communities fragmented by agricultural activities were analyzed in a 250-ha
area in eastern plains of Montecaseros, Gral. San Martín Department, Mendoza,
Argentina. A phytosociological method assessed different sites along a gradient
of human intervention, from natural environments with no evidence of altered
native vegetation to maximum farming modification, also including cleared and
abandoned fields. Soil analyses supplemented the characterization of six plant
communities. A scrubland physiognomy dominates the area, with species of the
genera Larrea, Atriplex and Lycium. Tillage and crop
abandonment can alter natural factors involved in soil formation, causing
deterioration and exerting selective pressure on species colonizing these
degraded environments. Evaluating natural vegetation before land clearing for
agriculture is essential to assess, through species that indicate environmental
conditions, edaphic limitations hindering crop establishment and affecting
productivity. The conservation of natural communities on private lands destined
for agriculture is valued.
Keywords: plant communities,
abandoned crops, dynamism, habitat fragmentation, soil degradation
Resumen
Se analizaron
comunidades vegetales fragmentadas por actividades agrícolas en un área de 250
ha en las llanuras orientales de Montecaseros, Departamento de Gral. San
Martín, Mendoza, Argentina. Se aplicó el método fitosociológico en sitios a lo
largo de un gradiente de intervención humana, desde ambientes naturales sin
evidencia de vegetación nativa alterada hasta la máxima modificación debido a
la agricultura, incluyendo también parcelas desmontadas y abandonadas. La
caracterización de seis comunidades vegetales se complementó con análisis de
suelo. La fisonomía dominante en la zona es el matorral, con especies de los
géneros Larrea, Atriplex y Lycium. La labranza y el
abandono de cultivos pueden alterar los factores naturales involucrados en la
formación del suelo. Estas actividades causan deterioro y ejercen una presión
selectiva sobre las especies que colonizan estos ambientes degradados. La
evaluación de la vegetación natural antes del desmonte para la agricultura es
fundamental para valorar, a través de especies indicadoras de las condiciones
ambientales, las limitaciones edáficas que dificultan el establecimiento de
cultivos y afectan su productividad. Se pone en valor la conservación de
comunidades naturales en terrenos privados destinados a la agricultura.
Palabras clave: comunidades
vegetales, cultivos abandonados, dinamismo, fragmentación de hábitat, degradación de
suelos
Originales: Recepción: 10/06/2022 - Aceptación: 11/12/2023
Introduction
Plant
biodiversity plays a strategic role in ecosystemic provision of goods and
services for human health and well-being (21).
Agriculture causes changes in these ecosystems involving both productive
activities and consequent effects on the socioeconomic and biophysical
environment. Agriculture requires ecosystem services (water, soil and genetic
diversity) while generating food, fiber and fuel, soil fertility, habitats for
wildlife, landscape and culture, among others (29).
However, the dominant models of agricultural intensification affect natural
resources by generating pollution, fragmentation and degradation, associated
with loss of ecosystem services (33).
Fragmentation of
natural environments consists of disintegrating a continuous landscape into
smaller fractions, separated from one another by a cover matrix resulting from
human activity (9). Replacement of
natural ecosystems for agroecosystems, urbanization, and other infrastructures
increases land vulnerability. Landscape fragmentation levels could be
considered an indicator of soil degradation (28)
generally related to inappropriate agricultural practices, overgrazing, unplanned
urbanization and expansion of agricultura frontier.
In Argentina,
66% of the territory corresponds to arid, semiarid and dry sub-humid areas,
where water and wind erosion affect two million hectares a year (16). In turn, about 47% of agricultural production
comes from drylands under desertification processes (2).
These ecosystems have low capacity for natural regeneration. Degradation
associated with reduced biological and economic productivity is hard to
reverse. Consequently, water and food supplies are affected, threatening
investment and technical development, and causing poverty and rural desertion,
feeding back the originating processes (27).
Mendoza plains,
where the most important agricultural activities take place, belongs to the
Monte Phytogeographic Province, an extensive vegetation unit that occupies
great part of Argentina’s drylands (7).
Given water deficit is the major factor limiting productivity, agriculture is
intensive and irrigated. Productive activities follow an agro-industrial model
in the large irrigated oases of Mendoza, already expanded over non-irrigated
areas. In San Martín (east Mendoza), more than 2200 farms comprise 94000 ha of
private producers, most of them landowners (14).
In Montecaseros, a district of San Martín, viticulture occupies most
agricultural land, where clearing has resulted in a mosaic of landscapes.
Productive crops merge with abandoned crops (with virtually bare soils,
evidence of degradation and low floristic diversity) and fragments of natural
fields (with appropriate native flora coverage preventing erosion processes).
Vegetation is an important component of landscape affected by geomorphology,
soil, climate, and even land use history (35).
Physiognomically, the area reflects environmental heterogeneity. Prosopis
flexuosa DC. woods and other areas representative
of the Monte biodiversity in Montecaseros provide fundamental ecosystem
services, where native plants might constitute reservoirs for pest predators
and crop pollinators.
At local level,
no publications addressing fragmentation in agricultural environments or
floristic-ecological studies are available. Related references in the region
involve ecology, management and conservation of Prosopis woodland (36), flora and vegetation in protected areas (25), vegetation assessment of peripheral oasis
lands (11), and vegetation coverage
classification in San Martín (19).
Considering
background information scarcity in San Martín and the importance of viticulture
for economic and social development, this study aimed to characterize plant
communities in natural and intervened environments, in a representative area of
the fragmentation processes caused by agriculture in Montecaseros district.
Materials
and methods
Study
area
The area
comprises 250 ha in Montecaseros district, General San Martín Department,
Mendoza, Argentina (figure 1), located on the eastern
fluvial-eolian plain of Mendoza (1),
concentrating the greatest agricultural activity of the province. Altitude
ranges between 598 and 644 m a. s. l. Climate is desertic (17), with mean annual temperature of 15°C, mean
annual rainfall reaching 200 mm under a spring-summer regime, and prevalence of
SE winds, frequently warm and dry (föehn type or “Zonda” winds).
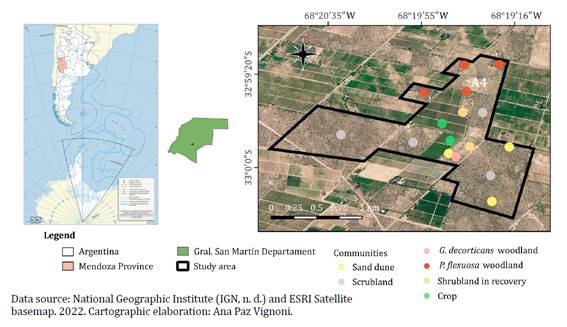
Figure 1. Study
area and distribution of communities surveyed.
Figura 1. Área
de estudio y distribución de las comunidades relevadas.
The vegetation
corresponds to the Monte Phytogeographic Province (7),
typical of the Northern District and Central Sub-district of the Monte Desert (26), recently considered within the Septentrional
District according to the latest classification (3).
In natural environments, evergreen shrub-steppe and scrubland are dominant,
represented by species of the genera Larrea, Atriplex and Lycium,
and small relics of Prosopis flexuosa and Geoffroea decorticans woodland
(Gillies ex Hook. & Arn.) Burkart. Grasses develop on open sites with low
tree and shrub coverage. Fixed and semi-fixed sand dunes are frequent, with
bare-root growth and psammophilous plants. Crops have fragmented these natural
vegetation areas, creating ideal conditions for the development of native and
exotic weed species.
Montecaseros
soils are poorly developed Entisols (23, 32)
with strong moisture déficit most of the year. Sandy textures prevail, with
surface or subsurface sites with finer soil layers, suitable for cropping given
groundwater availability. The entire area is affected by land degradation
processes, accelerated by natural water and wind erosion events.
Methodology
Analysis
of plant communities
Areas were
selected by satellite imagery (Google Earth, 2012) and available maps,
representing regions affected by fragmentation processes (figure
1). Research activities were conducted between December 2011 and June 2012.
Maps were made using free access QGIS software, version
3.22.6 (2022).
Plant
communities were studied by the phytosociological method (5), based on criteria of floristically,
physiognomically and ecologically homogeneous sites. Each species was assigned
abundance-dominance and sociability values in every location (24). In total, 87 surveys were conducted, and the
study area was delimited in the field according to community composition and
physiognomy. All plots were 49 m2 except at crop sites (30 m2) and in P.
flexuosa woodland, where surveys were done over a larger area (225 m2).
Additionally, each survey included vegetation stratification, geographic and
altitude coordinates (GPS) and visual assessment of slope decline, water of
dynamic and wind processes, solar exposure and evidence of human activity. Data
were first ordered and analyzed on a comparative table of surveys, and
summarized in a synthetic comparative table (table 1),
grouping those species best adjusted to each environment.
Table
1. Survey synthetic comparative table (24).
Tabla 1. Cuadro
comparativo sintético de relevamientos (24).
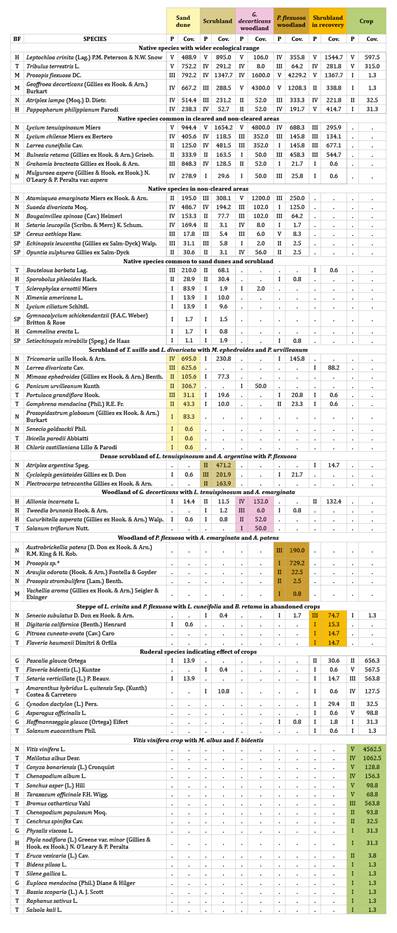
BF: biological forms (SP: succulent phanerophytes,
G: geophytes, H: hemicryptophytes, M: microphanerophytes, N: nanophanerophytes
and T: therophytes). P: species degree of presence (V: presence>80% of
surveys; IV: 60.1-80%; III: 40.1-60%; II: 20.1-40%; I: <20%). Cov.: global
coverage value (sum of mean species covers divided by total surveys per
community, multiplied by 100). Annex 2 details species with low values of
presence and/or coverage in each community. * Possible hybrid between P.
flexuosa and P. chilensis.
BF: formas biológicas (SP: fanerófitas suculentas,
G: geófitas, H: hemicriptófitas, M: microfanerófitas, N: nanofanerófitas y T:
terófitas). P: grados de presencia de cada especie (V: presencia >80% de los
relevamientos; IV: 60.1-80%; III: 40.1-60%; II: 20.1-40%; I: <20%). Cov.:
valor global de cobertura (suma de las coberturas medias de cada especie dividido el número total de los relevamientos de cada
comunidad por 100). El Anexo 2 detalla las especies con valores escasos de
presencia y/o cobertura en cada comunidad. * Posible híbrido entre P.
flexuosa y P. chilensis.
Categories
describing each group of species (24)
were 1- Exclusive characteristic species (strongly linked to a community,
practically not observed outside) or Preferential characteristic species (with
a wider ecological range but finding an optimum in one community), 2-
Accompanying species (wide ecological range, not related to a particular
community, indifferent) and 3- Accidental species (intruders in a community).
Floristic compositions of non-cleared sites (scrubland) were interpreted as
reference to the original natural vegetation. Table 2
synthesizes total mean coverage of each community, floristic richness and
dominant processes.
Table
2. Dominant processes and characteristics
of identified communities.
Tabla 2. Procesos
dominantes y características de las comunidades identificadas.
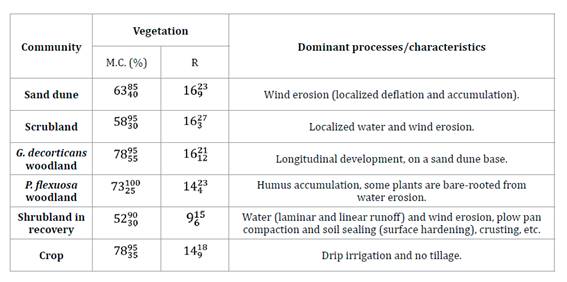
Total mean coverage (M.C.) expresses mean percentage
of soil covered with vegetation. Richness (R) represents mean number of species
per survey. Superindices and sub-indices show maximum and minimum observed values,
respectively.
La cobertura total media (M.C.) expresa el
porcentaje promedio de suelo cubierto por vegetación. La riqueza (R) representa
el número medio de especies por relevamiento. Superíndices y subíndices
muestran los valores máximos y mínimos observados, respectivamente.
Life form was
analyzed for each species according to perpetuation organ position (22) as succulent phanerophytes or cacti (SP,
plants with thick juicy stems), geophytes (G, plants with underground survival
organs), hemicryptophytes (H, herbs and perennial grasses with renewal buds at
ground level), microphanerophytes (M, trees with renewal shoots from 2 to 8 m),
nanophanerophytes (N, shrubs with renewal buds at less than 2 m height), and
therophytes (T, annual plants). In each community, relative contributions of
every biological form were estimated according to coverage and frequency, and
expressed in histograms or biological spectra (figure 2).
Coverage spectrums were determined after mean coverage values per
abundance-dominance interval of each species (24).
A frequency spectrum considered the number of species of each biological form
over total species in the community.
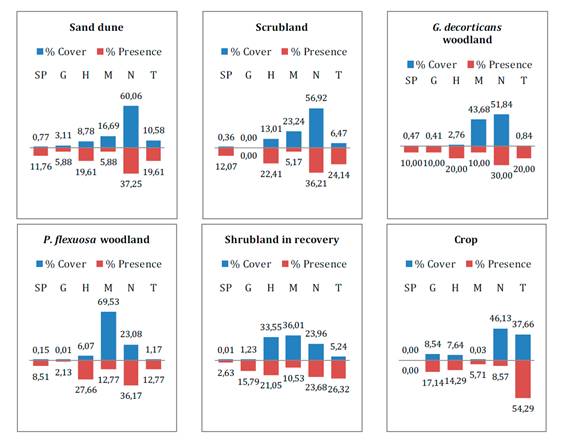
SP: succulent phanerophytes, G: geophytes, H:
hemicryptophytes, M: microphanerophytes, N: nanophanerophytes and T:
therophytes.
SP: fanerófitas suculentas, G: geófitas, H:
hemicriptófitas, M: microfanerófitas, N: nanofanerófitas y T: terófitas.
Figure 2. Biological
cover and frequency spectra for each community.
Figura 2. Espectros
biológicos de cobertura y frecuencia para cada comunidad.
Taxonomic
determinations
Representative
samples of species found in the area were herborized and taxonomically
identified based on reference specimens from the Mendoza Ruiz Leal Herbarium
(MERL, Argentine Institute for Dryland Research), and monographic works and
publications on Argentina’s regional floras. Regarding nomenclature, we
consulted the Catalog of Vascular Plants of Argentina (38) and the database of Argentine Flora (n. d.),
the International Plant Names Index (n. d.), Tropics organization (n. d.), and
the International Code of Nomenclature for algae, fungi, and plants (2018).
Recently, a disintegration of the genus Prosopis L. based on molecular
phylogeny has been proposed, followed by a new nomenclatural combination for
American species of the genera Neltuma and Strombocarpa (12). Nevertheless, we keep Burkart’s traditional
classification of Prosopis (6) for
nomenclatural stability, prevailing current use and disadvantageous changes
(31, articles 14, 34 and 56), until acceptance of nomenclatural changes by the
International Committee of Nomenclature.
Soil
analysis
Soil samples
supplemented environmental descriptions of different communities. After
establishing 16 sites, three samples were removed from a pit dug in each site,
achieving a maximum depth of 1.10 m, totalling 48 samples and including data
from the intermediate layer (0.20 to 0.50 m), where most root activity is
concentrated (table 3, Annex 1). Texture, fertility
(nitrogen, phosphorus, potassium and organic matter), salinity (electric
conductivity and sodium adsorption ratio), acidity (pH), ion complex (sodium,
calcium, magnesium, carbonate, bicarbonate, sulfate and chloride) and humidity
were determined for each simple (4).
Results were analyzed with descriptive statistics by InfoStat software version
2020 (8), graphed on star charts for
every community, where each variable is represented by a radius whose length is
the mean value of samples taken from each community (figure 3).
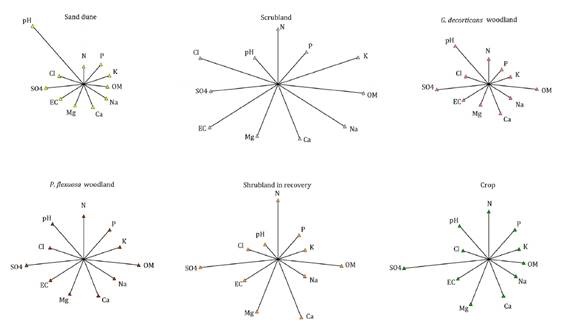
Each graph shows mean values of N: nitrogen, P:
phosphorus, K: potassium, OM: organic matter, Na: sodium, Ca: calcium, Mg:
magnesium, EC: electric conductivity, SO4: sulfates, Cl: chlorides and pH:
hydrogen potential.
En cada uno de los gráficos valores medios de N:
nitrógeno, P: fósforo, K: potasio, OM: materia orgánica, Na: sodio, Ca: calcio,
Mg: magnesio, EC: conductividad eléctrica, SO4: sulfatos, Cl: cloruros y pH:
potencial hidrógeno.
Figure 3. Star
graphs for main edaphic variables analyzed in each community.
Figura 3. Gráficos
de estrellas para las principales variables edáficas analizadas en cada
comunidad.
Results
and discussion
Six plant
communities were identified as sand dunes, scrubland, G. decorticans woodland,
P. flexuosa woodland, shrubland in recovery after crop abandonment, and
crop (photo 1; table 1; figure
1). Species with wider ecological range (accompanying species) were L.
crinita, T. terrestris, P. flexuosa, G. decorticans,
A. lampa and P. philippianum.

Photo 1. Study
area: A, crop, B, shrubland in recovery and C, non-cleared scrubland.
Foto 1. Área
de estudio: A, cultivo, B, arbustal en recuperación y C, matorral sin
desmontar.
Poor soils in
organic matter and nitrogen had good contents of phosphorus and potassium, and
different levels of salinity. Most of the samples showed mildly alkaline and
non-sodic characteristics (table 3, Annex 1).
Scrubland
of T. usillo and L. divaricata with M. ephedroides
and P. urvilleanum on sand dunes
Sand dunes had
predominant scrub vegetation and small isolated trees, distributed in three
vegetation-height layers as trees dominated by P. flexuosa and G.
decorticans (average height of 3 m), shrubs dominated by T. usillo and
L. divaricata (1.5 m), and an herbaceous layer dominated by L.
crinita and P. urvilleanum (0.5 m). Heterogeneous sun exposure of
this natural topography (from 5 to 10 m in height) was evidenced in vegetation
growth. Thus, P. urvilleanum forms compact colonies and is occasionally
dominant in dry sunny áreas (fixing and stabilizing more exposed dune sectors),
whereas Z. jamesonii grows mostly in shady places, reflecting the
micro-environmental diversity of dunes and being one of the greatest floristic
resources of the study area (table 2).
Exclusive
characteristic species of this community resulted P. urvilleanum, S.
goldsackii, P. globosum, I. parodii and C. castilloniana,
preferential characteristic species are T. usillo, M. ephedroides,
L. divaricata, P. grandiflora, G. mendocina, X.
americana and T. terrestris, and accompanying species are
represented by A. lampa, L. chilense, M. aspera, B.
spinosa, S. divaricata, G. bracteata, S. leucopila, B.
barbata, S. arnottii and E. leucantha.
Species coverage
and presence spectra reflected a community dominated by nanophanerophytes
(distinctive plants in a sand dune physiognomy), therophytes and
hemicryptophytes. Microphanerophytes, although not frequent, seemed to
contribute considerable coverage in contrast to the numerous succulent
phanerophytes, of insignificant relative coverage (figure 2).
Sand dunes are generally not saline due to
soil permeability. The low water content of soil surface layers hinders organic
matter decomposition and nutrient fixing, resulting in low fertility.
Nevertheless, higher nitrogen values were found on shady sides of the dune,
closest to crops and coinciding with a higher density of P. flexuosa trees,
harbouring atmospheric nitrogen-fixing bacteria in their roots (34). Dunes pH was moderately to strongly
alkaline, consistent with higher carbonate and bicarbonate values.
Dense
scrubland of L. tenuispinosum and A. argentina with P. flexuosa
This community,
on sites with no evidence of clearing, is mainly composed of tall shrubs and
trees, isolated or in small groups (photo 2). It presented
three height layers, tres dominated by P. flexuosa (average height of 3
m), shrubs dominated by L. tenuispinosum, L. cuneifolia and A.
argentina (1.8 m), and an herbaceous layer dominated by L. crinita (0.5
m). These scrublands on saline soils occupy the greatest land expanse in the
study area, fragmented by vineyards. Tall and dense scrubland is
physiognomically predominant in the landscape, alternating with sparse
shrubland, sandy deposit sites, ground surface wáter accumulation, or evidence
of laminar runoff in natural drainage areas. This scrubland would be considered
a reference of typical natural vegetation in the area, helping understand plant
plot recovery after crop abandonment, since both communities show similar
edaphic and floristic features.

Photo 2. Overall
scrubland physiognomy.
Foto 2. Fisonomía
general del matorral.
Exclusive
characteristic species are P. tetracantha, G. boliviana and M.
ovata in Sandy soils, and A. spegazzinii in saline, finer-textured
soils. All preferential characteristic species like A. argentina, C.
genistoides, S. mirabilis and S. phleoides are typical of
saline soils. Accompanying species are P. flexuosa, A. emarginata and
T. usillo in sandy areas, and L. tenuispinosum, L. crinita,
L. cuneifolia and S. divaricata in saline or floodable lands. A.
hybridus and F. bidentis appear as accidental species after nearby
crop influence.
Scrubland
physiognomy was evidenced by dominance of nanophanerophytes, followed by
microphanerophytes. Therophytes and hemicryptophytes species were abundant. All
succulent phanerophyte species surveyed in this study are represented in this
community. Scrubland soil varied depending on the area surveyed, with sandy to
loamy textures, low fertility, on slightly to excessively saline sites. In the
latter case, the highest soluble sodium salt content (chlorides and sulfates)
indicated strongly sodic soils.
Closed
dune-base woodland of G. decorticans with L. tenuispinosum and A.
emarginata
Closed G.
decorticans woodland with three height layers included trees dominated by G.
decorticans with some P. flexuosa trees (heights from 1.5 to 5 m),
shrubs dominated by L. tenuispinosum and A. emarginata (average
height of 1 m), and an herbaceous layer with low coverage values, dominated by L.
crinita (0.80 m) and A. incarnata (0.1 m). Geoffroea decorticans woodland,
longwise developed, was found at the base of a sand dune, spreading by
gemmiferous roots, facilitating colonization of the abandoned crop area in recovery
process.
Solanum
triflorum was
found exclusive to this community, beneath P. flexuosa canopies. Preferential
characteristic species are G. decorticans, L. tenuispinosum and O.
sulphurea (tolerant to floodable, clay or saline soils); A. emarginata and
T. brunonis (representative of P. flexuosa woodland), and A.
incarnata and C. asperata (linked to sandy soils by proximity to the
sand dune of G. decorticans woodland). Accompanying species observed included
P. flexuosa, L. cuneifolia and L. chilense, notoriously
affecting total coverage and presence, where 95% of the relative cover is
contributed by nanophanerophytes and microphanerophytes, and 4% by
hemicryptophytes and therophytes. This is the only community comprising all
life forms, showing a more equitable distribution of presence values.
Soil is
sandy-loam up to 0.70 m deep, and sandy at greater depths. Fertility is low and
the sand dune base determines good water drainage, reflected in the
low-salinity soil profile.
Woodland
of P. flexuosa with A. emarginata and A. patens
Relics of P.
flexuosa woodland, fragmented by crops or cleared areas, were surveyed on
12 plots of 225 m2 each showing open P. flexuosa groves (trees between
2.5 and 5 m tall, whose canopies rarely touch), or closed and semi-closed
groves (trees up to 6.5 m tall, whose canopies frequently touch). Three height
layers were distinguished with tres dominated by P. flexuosa, sometimes
accompanied by G. decorticans and B. retama (average height of 3
m), shrubs dominated by L. tenuispinosum and/or A. emarginata (1
m), and an herbaceous layer dominated by L. crinita (0.80 m). We
observed P. flexuosa trees with a single trunk and multiple stems and
mean tree coverage ranging between 22.5% and 62.5% (value explained by the
great number of saplings with main shoot diameter under 5 cm: 130/survey, on
average).
A. patens, P.
strombulifera, A. odorata, V. astringens and V. aroma are
exclusive characteristic species in this community. P. flexuosa reaches
the highest relative coverage in the area, as well as A. mendocina and S.
aphylla, identified as preferential characteristic species. P. flexuosa canopies
prevent light from reaching the ground, limiting development of succulent
phanerophytes, geophytes and therophytes with low relative coverage values.
Soil samples
were collected from the grove adjoining the crop (A4, figure 1), finding non-saline
soils of sandy loam texture up to 0.20 m deep and sandy at greater depths. Nitrogen
and organic matter contents were among the highest recorded at all sites in the
study area and, just as phosphorus and potassium contents, reached higher
values in surface layers, probably induced by humus beneath tree canopies (34).
Low
and open steppe of L. crinita and P. flexuosa with L.
cuneifolia and B. retama in abandoned crops
Open steppe (52%
of mean total coverage) showed shrubs and grasses with young, isolated, small,
multi-stemmed trees (photo 3).

Photo 3. Physiognomy
of shrubland under recovery, colonized by grasses. Old crop furrows with
evidence of water erosion are observed.
Foto 3. Fisonomía
del arbustal en recuperación, colonizado por gramíneas. Se observan los
antiguos surcos de cultivo con evidencias de erosión hídrica.
Three height
layers were distinguished by trees dominated by P. flexuosa and B.
retama (average height of 2 m), shrubs dominated by L. cuneifolia (1
m), and an herbaceous layer of L. crinita and P. philippianum (0.80
m). This community corresponds to cleared areas levelled in 1972 for crop
establishment (grapes, apricots, alfalfa and vegetables), labored for as long
as 15 years and abandoned in 1987 due to poor performance and yield.
Agricultural tillage altered soil characteristics in this area during 25 years
between abandonment and present surveys. The surrounding natural vegetation has
recolonized this site with numerous species, typical of fine-textured, saline or
floodable soils. Adjoining crops contribute to species surviving the edaphic
limitations of these abandoned plots.
Flaveria
haumanii and
P. cuneato-ovata behave as exclusive characteristic species; L.
crinita, P. philippianum, L. cuneifolia, B. retama, S.
subulatus and D. californica as preferential characteristic species,
depending on site propensity for salt and water accumulation, or surface runoff
after rainfall (25). P. flexuosa,
as an accompanying species, contributes important coverage and presence, and C.
dactylon and P. glauca, as accidental species, denote close
crop-area influence.
Hemicryptophytes
(primarily L. crinita) establishment increases in sunny areas with
almost non-competition, defining the vegetation physiognomy together with
nanophanerophytes and microphanerophytes. Scarce succulent phanerophytes and
geophytes were evidenced by these plant limitation to propagate from seeds or
in agamic reproduction.
Soils resulted
sandy loam, with nitrogen and organic matter contents similar to non-cleared
sites. Salinity increased with depth. A gypsum layer detected in the soil
profile was analytically confirmed by higher calcium sulfate concentrations.
This pattern may be due to machinery effects during years of tillage,
generating a deep compacted layer that favoured salt concentration in the soil
profile after periodic irrigation was suspended.
In general
terms, these results are consistent with other studies associating site use
history, particularly productive plots abandonment with higher soil erodibility
and changes in landscape physiognomy of the Monte native vegetation (35, 37).
Vitis
vinifera crop
with M. albus and F. bidentis
A 31-ha vineyard
with different grapevine cultivars, conducted both vertically and horizontally
under drip irrigation, presented two height layers, one shrub layer dominated by
V. vinifera (average height of 2 m) and a herbaceous layer dominated by M.
albus, F. bidentis, S. verticillata and B. catharticus (0.5
m). The vineyard was surrounded by cleared areas with spontaneously recovered
vegetation, and by other areas with undisturbed natural vegetation, a sand
dune, and P. flexuosa groves.
Exclusive
characteristic species with higher coverage values were V. vinifera, M.
albus, B. catharticus, C. album, C. bonariensis, S.
asper and T. officinale. Overall, they were widely distributed
(normally adventitious or introduced). Despite great multiplying capacities, these
species require more water than other drought-tolerant ones, such as H. glauca,
P. glauca, C. dactylon, F. bidentis, A. hybridus, S.
verticillata, A. officinalis and S. euacanthum, all preferential
characteristic vineyard species, although capable of surviving in less
favorable conditions. L. crinita (more abundant at sunlit crop sites as
row ends, borders, plots with young vines, with less coverage or lower height)
and T. terrestris behave as accompanying species, whereas P. flexuosa
and G. decorticans are accidental, with small presence, coming from
adjoining scrublands.
Dominant in
biological spectra are nanophanerophytes (primarily grapevines, contributing
nearly half the coverage despite low presence) and therophytes (generally small
and abundant). Geophytes reach the highest coverage and frequency values,
probably because tillage does not remove rhizomes, bulbils or other organs of
asexual reproduction. Low microphanerophytes frequency is explained by spontaneous
P. flexuosa and G. decorticans individuals.
Soils are sandy
on the surface and intermediate layers and sandy -loam at depths greater than
0.5 m, with low nitrogen and organic matter contents, non to slightly saline.
Conclusions
Scrubland is the
predominant physiognomy in the area, typical for the Monte Province, with
species of the genera Larrea, Atriplex and Lycium. According
to this study, the prevailing vegetation before crop establishment was
halophilic or salt-tolerant. Species like A. lampa, A. spegazzinii,
A. argentina, P. tetracantha, G. bracteata and S.
divaricata are frequent and grow in almost all communities identified.
Prosopis
flexuosa groves
grow where trees can benefit from groundwater, bordering the steppe or in inter-dune
areas. As woodland canopy density increases, lower-height vegetation layers
tend to diminish and even disappear due to sunlight shortage, except for
sciophilous species like A. emarginata. G. decorticans is
abundant on sites with finer-textured soil, at the base of sand dunes or in
depressed areas, growing in well-defined groves. Through powerful gemmiferous
roots, this species is one main colonizer of abandoned crop fields.
In sand dunes
and their surroundings, psammophilous vegetation is well represented by a great
diversity of species, given the micro-environmental features of these
formations. P. urvilleanum is especially frequent in sand dunes
destabilized by anthropic action. Adventitious or introduced species, dependent
on adequate water supply (M. albus, C. album, S. asper, T.
officinale, etc.) coexist in combination with more rustic species able to
develop under water and/or saline stress (F. bidentis, P. glauca,
A. hybridus, H. glauca, S. verticillata, C. dactylon,
etc.). Thus, a crop area is the source of propagules dispersal for these last
species being introduced into neighbouring sites.
In shrubland
under recovery, clearing, tillage and later crop abandonment emphasized edaphic
constraints, analytically determined by signs of soil erosion, compaction or
crusting. Spontaneous re-vegetation led to lower floristic richness compared to
the adjacent natural areas taken as reference sites, after 25 years of
abandonment, with strong presence of tolerant species or species indicating
fine-textured soils, floodable areas, or with surface runoff and saline
characteristics. Original shrubland has turned into a steppe of bushes
(halophilic in part) and grasses with small young trees. Several species
present in the surroundings (O. sulphurea, S. divaricata, S.
mirabilis, C. genistoides and P. tetracantha, among others)
had difficult establishments on the plot, despite being halophilic or
salt-tolerant. This result supports the hypothesis that the main problem for
plot recovery would not be saline content, but soil structure alteration by
tillage and subsequent crop abandonment, reinforcing uncertainty about
biodiversity restoration levels to be sought in degraded soils, and recovery
time span. This study demonstrated that a survey of natural environments with
proper assessment methodologies is fundamental before land clearing for
agriculture. Determination of plant communities in fragmented areas, and
valuation of species indicative of edaphic limitations restricting crop
establishment and productivity, are essential in defining land uses.
Flora components
and species distribution are adequate indicators for monitoring environmental
processes in plant communities. Incorporating the private sector into
environmental protection systems towards the establishment of isolated
preserved sites is necessary but not sufficient in disjointed landscapes. This
first analysis of natural áreas fragmented by agriculture in Montecaseros,
shows that typical communities of the Monte subsist, demonstrating how diverse
flora is preserved in privately owned lands. Changes in floristic composition
of plant communities may have occurred in the study area during the time
elapsed between this survey and the current situation. The results obtained
constitute an important baseline for monitoring sampling sites and vegetation
dynamics in fragmented areas of Montecaseros. Cultivable land is a vulnerable
and scarce resource, particularly in drylands. A proper interpretation
recognizes limiting territory factors while approaching systemic criteria
highlighting complementarity in management between crops and native vegetation
areas. Studies of vegetation dynamism in private lands will contribute to plan
and implement sustainable land management, tending to recover and conserve
biodiversity in degraded soils, considering all associated ecosystem services.
Acknowledgements
We thank María
Margarita González Loyarte for her accurate observations and valuable review; Pablo
Molina for his generous collaboration in field work and taxonomic determinations,
and Eliana Vargas for her advice in statistic data analysis.
1. Abraham, E.
M. 2000. Geomorfología de la Provincia de Mendoza. Argentina. Recursos y
problemas ambientales de la zona árida. Provincias de Mendoza, San Juan y La Rioja.
1: 29-48.
2. Abraham, E.;
Rubio, C.; Salomón, M.; Soria, D. 2014. Desertificación: problema ambiental
complejo de las tierras secas. Ventanas sobre el territorio. Herramientas
teóricas para comprender las tierras secas. Mendoza: EDIUNC. 187-265.
3. Arana, M. D.;
Natale, E. S.; Ferretti, N. E.; Romano, G. M.; Oggero, A. J.; Martínez, G.;
Posadas, P. E.; Morrone, J. J. 2021. Esquema biogeográfico de la República
Argentina. Opera Lilloana. 56: 238.
4. Association
of Official Analytical Chemists (AOAC). 2019. International, Official Methods
of Analysis, 21st ed., Gaithersburg, USA. URL: https://www.aoac.org/resources
5.
Braun-Blanquet, J. 1979. Fitosociología. Bases para el estudio de las
comunidades vegetales. H. Blume. Madrid.
6. Burkart, A.
1976. A monograph of the genus Prosopis (Leguminosae subfam. Mimosoideae).
Journal of the Arnold arboretum. 57: 219-249 and 450-525.
7. Cabrera, A.
1971. Fitogeografía de la República Argentina. Boletín de la Sociedad Argentina
de Botánica. Volumen XIV. (1-2).
8. Di Rienzo, J.
A.; Casanoves, F.; Balzarini, M. G.; González, L.; Tablada, M.; Robledo, C. W.
2020. InfoStat, versión 2020. Centro de Transferencia InfoStat, Universidad
Nacional de Córdoba, Argentina. URL: http://www.infostat.com.ar
9. Fahrig, L.
2003. Effects of habitat fragmentation on biodiversity. Annual review of
ecology, evolution, and systematics. 34(1): 487-515.
10. Flora
Argentina y del Cono Sur. Flora vascular de la República Argentina y del Cono
Sur. N.d. http://www.floraargentina.edu.ar/
11. González
Loyarte, M. M.; Gaviola, S.; Buk, E.; Rodeghiero, A. G.; Menenti, M. 2007.
Propuesta metodológica para la evaluación de tierras periféricas al oasis
irrigado de Lavalle, Mendoza (Argentina). Revista de la Facultad de Ciencias
Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. 39(2): 109-126.
12. Hughes, C.
E.; Ringelberg, J. J.; Lewis, G. P.; Catalano, S. A. 2022. Disintegration of
the genus Prosopis L. (Leguminosae, Caesalpinioideae,
mimosoid clade). PhytoKeys. 205: 147-189. https://doi.org/10.3897/phytokeys.205.75379
13. Instituto
Geográfico Nacional (IGN). N.d. Información geoespacial. Accessed 02.05.22.
https://www.ign.gob.ar/
14. Instituto
Nacional de Estadística y Censos (INDEC). 2021. Censo Nacional Agropecuario
2018: resultados definitivos. Ciudad Autónoma de Buenos Aires.
15. IPNI. 2023.
International Plant Names Index. Published on the Internet http://www.ipni.org, The Royal Botanic Gardens, Kew, Harvard
University Herbaria & Libraries and Australian National Herbarium.
16. Ministerio
de Ambiente y Desarrollo Sostenible de la Nación (MAyDS). 2020. República
Argentina. Reporte final sobre el Programa de Establecimiento de Metas
Voluntarias de Neutralidad de la Degradación de la Tierra. Ciudad de Buenos
Aires. https://www.argentina.gob. ar/ambiente/contenidos/faros-de-conservacion
17. Norte, F.
2000. Mapa climático de Mendoza. Argentina. Recursos y Problemas Ambientales de
la Zona Árida. Primera Parte. Provincias de Mendoza, San Juan y La Rioja. 1:
25-27.
18. Pérez
Valenzuela, B. R. 1999. Edafología en la agricultura regadía cuyana. Fundar.
205 p.
19. Pérez
Valenzuela, B. R.; Salcedo, C. E.; De Cara, D. E.; Capuccino, S. N. 2001.
Vegetación de San Martín (Mendoza). Revista de la Facultad de Ciencias Agrarias .
Universidad Nacional de Cuyo. Mendoza. Argentina. 33(2): 63-71.
20. QGIS
Development Team. 2022. QGIS Geographic Information System. Open Source
Geospatial Foundation Project. https://qgis.org/es/site/
21. Quijas, S.;
Schmid, B.; Balvanera, P. 2010. Plant diversity enhances provision of ecosystem
services: A new synthesis. Basic and Applied Ecology. 11(7): 582-593.
https://doi.org/10.1016/j.baae.2010.06.009
22. Raunkiaer,
C. 1934. Life forms and terrestrial plant geography. Clarendon Press, Oxford.
23. Regairaz, M.
C. 2000. Suelos de Mendoza. Argentina. Recurso y problemas ambientales de la
zona árida. Provincia de Mendoza, San Juan y La Rioja. 1: 59-62.
24. Roig, F. A.
1973. El cuadro fitosociológico en el estudio de la vegetación. Deserta. 4:
45-67.
25. Roig, F. A.
1981. Flora de la reserva ecológica de Ñacuñán. Cuaderno Técnico IADIZA. 3(80):
5-176.
26. Roig, F. A.;
Roig-Juñent, S.; Corbalán, V. 2009. Biogeography of the Monte desert. Journal
of Arid Environments. 73(2): 164-172.
https://doi.org/10.1016/j.jaridenv.2008.07.016
27. Salvia, R.;
Quaranta, V.; Sateriano, A.; Quaranta, G. 2022. Land Resource Depletion,
Regional Disparities, and the Claim for a Renewed ‘Sustainability Thinking’
under Early Desertification Conditions. Resources. 11(3): 28.
https://doi.org/10.3390/resources11030028
28. Smiraglia,
D.; Tombolini, I.; Canfora, L.; Bajocco, S.; Perini, L.; Salvati, L. 2019. The
latent relationship between soil vulnerability to degradation and land
fragmentation: A statistical analysis of landscape metrics in Italy, 1960-2010.
Environmental management. 64(2): 154-165. https://doi.org/10.1007/s00267-019-01175-6
29. Swinton, S.
M.; Lupi, F.; Robertson, G. P.; Hamilton, S. K. 2007. Ecosystem services and
agriculture: cultivating agricultural ecosystems for diverse benefits. Ecological
economics. 64(2): 245-252. https://doi.org/10.1016/j.ecolecon.2007.09.020
30.
Tropicos.org. Missouri Botanical Garden. https://tropicos.org
31. Turland, N.
J.; Wiersema, J. H.; Barrie, F. R.; Greuter, W.; Hawksworth, D. L.; Herendeen,
P. S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; May, T. W.; McNeill,
J.; Monro, A. M.; Prado, J.; Price, M. J.; Smith, G. F. (eds.) 2018.
International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code)
adopted by the Nineteenth International Botanical Congress Shenzhen, China,
July 2017. Regnum Vegetabile 159. Glashütten: Koeltz Botanical Books. DOI
https://doi.org/10.12705/Code.2018
32. United
States Department of Agriculture (USDA). 1954. Diagnosis and improvement of
saline and alkali soils. Agriculture Handbook 60, Richards, L.A. (Ed.). U. S.
Laboratory Staff, Washington, DC.
33. Viglizzo, E.
F.; Frank, F. C.; Carreño, L. V.; Jobbágy, E. G.; Pereyra, H.; Clatt, J.;
Pincen, D.; Ricard, M. F. 2011. Ecological and environmental footprint of 50 years
of agricultural expansion in Argentina. Global change biology, 17(2): 959-973. doi: 10.1111/j.1365-2486.2010.02293.x
34. Villagra, P.
E. 2000. Aspectos ecológicos de los algarrobales argentinos. Multequina. 9(2):
35-51.
35. Villagra, P.
E.; Defossé, G. E.; Del Valle, H. F.; Tabeni, S.; Rostagno, M.; Cesca, E.;
Abraham, E. 2009. Land use and disturbance effects on the dynamics of natural
ecosystems of the Monte Desert: Implications for their management. Journal of
Arid Environments. 73(2): 202-211. https://doi.org/10.1016/j.jaridenv.2008.08.002
36. Villagra, P.
E.; Álvarez, J. A. 2019. Determinantes ambientales y desafíos para el
ordenamiento forestal sustentable en los algarrobales del Monte, Argentina.
Ecología Austral. 29(1): 146-155. https://doi.org/10.25260/EA.19.29.1.0.752
37. Yannelli, F.
A.; Tabeni, S.; Mastrantonio, L. E.; Vezzani, N. 2014. Assessing degradation of
abandoned farmlands for conservation of the Monte Desert biome in Argentina.
Environmental management. 53(1): 231-239.
38. Zuloaga, F.;
Morrone, O. 1999. Catálogo de las Plantas Vasculares de la República Argentina.
II. Monogr. Syst. Bot. Missouri Bot. Gard. 74.
https://drive.google.com/file/d/1ST-hgPbnmvltbARGf2o7KkoD0IXoNFGP/view?usp=sharing