Revista de la Facultad de Ciencias
Agrarias. Universidad Nacional de Cuyo. Tomo 56(1). ISSN (en línea) 1853-8665.
Año 2024.
Original article
Landform
heterogeneity drives multi-stemmed Neltuma flexuosa growth dynamics.
Implication for the Central Monte Desert forest management
La
heterogeneidad de paisaje modula la dinámica en el crecimiento de Neltuma
flexuosa. Implicancias para el manejo forestal de los bosques del Desierto
del Monte Central
1Laboratorio
de Dendrocronología e Historia Ambiental (IANIGLA CONICET). Av. Dr. Adrian Ruiz
Leal. Mendoza. M5500. Argentina.
2Universidad
Nacional de Cuyo. Facultad de Ciencias Agrarias. Cátedra de Dasonomía.
Almirante Brown 500 (M5528AHB). Chacras de Coria. Mendoza. Argentina.
3Universidad
Mayor. Facultad de Ciencias. Hémera Centro de Observación de la Tierra. Camino
La Pirámide 5750. Huechuraba. Santiago. Chile.
*spiraino@fca.uncu.edu.ar
Abstract
Drylands
represent the main earth biome, providing ecosytemic services to a large number
of people. Along these environments, woodlands are often dominated by
multi-stemmed trees, which are exploited by local inhabitants to obtain forest
products for their livelihood. In central-west Argentina, Neltuma flexuosa (algarrobo)
woodlands are distributed across different landform units, varying in
topographical and soil characteristics. This research aimed to reconstruct
stem-growth time until harvestable diameter was achieved, and biological
rotation age according to topo-edaphic variability in three algarrobo forests
using dendrochronological methods. Results indicated that landform heterogeneity
modulated species radial growth, influencing stem increments and cutting cycle
period. In this sense, a decreasing trend in tree productivity emerged along a
loamy-to-sandy textured soil gradient. These findings provide useful novel
information for N. flexuosa forest management, suggesting the need to
account for spatial landform/soil heterogeneity when examining desert forest
dynamics.
Keywords: arid woodlands,
forest management, growth form, tree-rings
Resumen
Las tierras
secas representan el principal bioma terrestre, proveyendo numerosos servicios
ecosistémicos a un ingente número de personas. En estos ambientes, los bosques
son a menudo dominados por árboles de porte multi-fustal, explotados por los
habitantes locales para obtener productos forestales útiles para su sustento.
En los territorios del centro-oeste de Argentina, los bosques de Neltuma
flexuosa (algarrobo) están distribuidos en diferentes unidades de paisaje,
los que presentan variaciones en las características topográficas y edáficas.
En este estudio reconstruimos, a través de métodos dendrocronológicos, el
crecimiento, el intervalo para obtener productos maderables, y el turno
biológico de corta en función de la variabilidad topo-edáfica en tres bosques
diferentes de algarrobo. Los resultados indican que la heterogeneidad del
paisaje modula el crecimiento radial de la especie, influenciando el incremento
diamétrico y el turno de corta. En este sentido, se evidencia una tendencia
decreciente en valores de productividad según un gradiente edáfico desde
textura limosa a arenosa. Estos hallazgos proveen información novedosa y de
utilidad para los planes de manejo forestal de los bosques de N. flexuosa,
indicando la necesidad por considerar la heterogeneidad
espacial establecidas por las unidades de paisaje/suelos en estudios
relativos a la dinámica forestal de los bosques de tierras secas.
Palabras claves:
bosques
de tierras secas, manejo forestal, forma de crecimiento, ancho de anillo
Originales: Recepción: 18/09/2023 - Aceptación: 14/04/2024
Introduction
Worldwide,
drylands represent the largest biome on Earth, covering more than 40% of land
surface (7, 29). These environments,
characterized by precipitation/evapotranspiration ratio below 0.2, include 20%
of plant diversity hotspots, play a fundamental role in various biogeochemical
cycles, and contribute to approximately 40% of global net primary productivity
(11, 16, 29).
Drylands host
more than two billion people (29). Along
these biomes, forests cover approximately 1.400 Mha, providing several
ecosystemic services like water and soil regulation, food, biochemical, and raw
material provision, as well as cultural services (7,
24). Physiognomically, many desert forests are dominated by tree species
capable of regeneration through development of new tissue following
disturbance, a physiological trait known as resprouting (8, 30). This results in vegetation with
coexisting one- and multi-stemmed trees, which differ in growth, productivity, reproduction,
and survival ability (30).
Dryland forests
have suffered various types of natural and anthropogenic disturbances, such as,
fire, insect outbreak, and drought, in addition to high deforestation rates due
to agricultural and livestock expansion (6, 20).
In this sense, it is estimated that, globally, about 220,000 km2 of
tree-covered drylands were converted into other land cover types between 1992
and 2015 (29). This suggests the need to
sustainably exploit and manage these natural resources, particularly in the
actual climate change scenario (16).
The Central
Monte Desert (hereafter, CMD) is a dryland biome located in central-west Argentina
(2). Along these districts, the algarrobo
dulce, Neltuma flexuosa (DC.) C.E. Hughes & G.P. Lewis (formerly designated
as Prosopis flexuosa DC), is a dominant tree species growing on a
variety of landform units (20). These
woodlands were severely exploited during the first half of the 20th
century, mainly for domestic wood demand, railway construction and, afterwards,
vineyard expansion, with a resulted extraction of approximately 1 million tons of
wood (1, 28).
Nowadays, the N.
flexuosa CMD forests are mostly managed by the local Huarpe native community,
through forestry practices that consider the typical low tree growth rates of these
woodlands (28). As a result, logging and
thinning are extremely rare activities in this area, whereas other
silvicultural approaches, such as pruning and deadwood removal, represent
valuable forest management alternatives (4, 27).
Pruning plays a prominent role due to the high presence of multi-stemmed trees
in CMD forests, as well as their higher productivity with respect to
one-stemmed individuals (3, 4).
Previous
analyses examined productivity in one- and multi-stemmed N. flexuosa trees,
providing valuable information regarding the CMD algarrobo forest management,
but solely at local scale (4). As
previously mentioned, the CMD N. flexuosa woodlands grow on a mosaic of
landform units, expression of soil variability and topographic characteristics
(21). This environmental heterogeneity is
reflected in differences of tree-ring development, as well as its relation with
precipitation regime and disturbance (21, 22).
It could be hypothesized, therefore, that the CMD landform variability would
represent a factor modulating multi-stemmed N. flexuosa tree
productivity, as well as stem-cutting cycle periods. Studying how topo-edaphic
characteristics influence multi-stemmed algarrobo growth dynamics can enhance
CMD silvicultural management by identifying landforms that can sustain intense
forest exploitation without ecological imbalance. For this reason, this
research analyzed stem growth rates of several multi-stemmed N. flexuosa trees
distributed along three different landform units, aiming to reconstruct
diametric cumulative increment, time (expressed in years) until forest products
can be obtained, and biological rotation age according to variations in
topography and soil characteristics of the CMD algarrobo forest stands.
Materials
and methods
In this study,
we examined three sites within the CMD located in the Province of Mendoza,
Argentina, representing distinct landform units (figure 1).
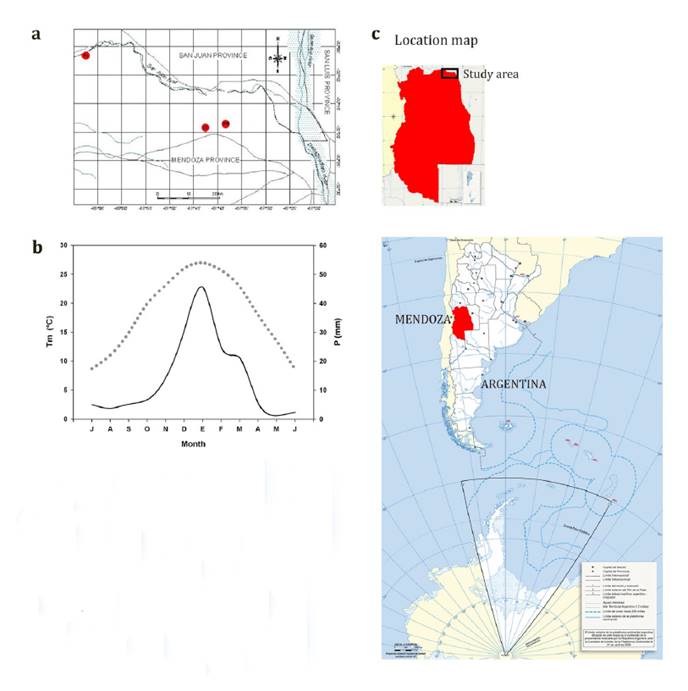
Monthly total precipitation (continuous line) and
monthly air temperature (dashed line) data belong to the Encón climatic station
(32°15’ S, 67°47’ W), covering the 1971-1987 period. RI, PR, and ID represent
River, Paleo-river, and Inter-dune valley units, respectively.
Los datos de precipitación media mensual (línea
continua) y temperatura promedio mensual (línea puntuada) pertenecen a la
estación meteorológica de Encón (32°15’ S, 67°47’ W), para el período
1971-1987. RI, PR, e ID representan respectivamente las unidades Riparia,
Paleo-riparia, y de Valle inter-medanoso.
Figure 1. Spatial
position of the sampled plots (a), climatic diagram (b), and geographical
location of the study area (c).
Figura 1. Ubicación
de los sitios de muestreo (a), diagrama climático (b), y localización
geográfica del área de estudio (c).
Along these
districts, the algarrobo woodlands grow under semi-arid climatic conditions,
with mean annual precipitation of 155 mm, and large variability in temperature
at both daily and seasonal timescales (2).
Vegetation consists of the typical CMD plant association, with N. flexousa dominating
the tree layer, occasionally accompanied by Geoffroea decorticans, along
with shrub presence of Larrea divaricata Cav., (Hook. And Arn.), Atamisquea
emarginata Miers ex Hook. & Arn and Bulnesia retama (Hook.)
Griseb (28).
Sites were selected
based on prior geomorphological classification (23).
In this study, River, Paleo-river, and Inter-dune valley landform units were
examined, differing in topographic characteristics and edaphic features (table 1).
Table
1. Geographical and environmental
characteristics of the sampled sites.
Tabla 1. Características
geográficas y ambientales de los sitios de muestreo.
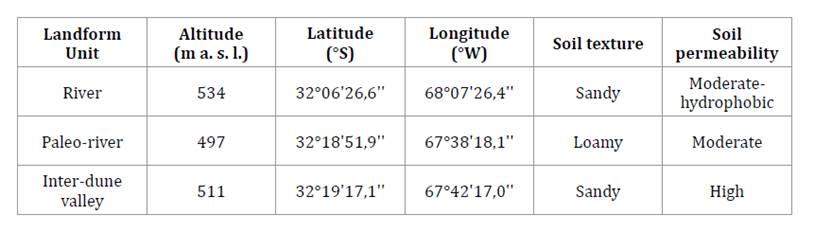
m
a. s. l.: meters above sea level.
m
a. s. l.: metros sobre el nivel del mar.
The selected
algarrobo woodlands were in relative proximity (within 50 km), allowing us to
assume a common climatic influence upon the species stem growth. Soil types
varied among the examined stands, with sandy soils in the River and Interdune
valley units and loamy soils in the Paleo-river site (table 1).
On the other hand, soil permeability differed among the selected woodlands,
showing high to moderate and moderate-hydrophobic characteristics (table
1).
At each site,
1-2 rectangular plots of 1.000m2 (50m x 20m) were established to
examine the algarrobo forest dynamics in terms of tree radial growth relation
with rainfall and its response to thinning, results already published (21, 22). The examined stands exhibited variations
in structural characteristics, including differences in multi-stemmed tree density,
mean basal stem diameter, and tree height, whereas the number of stems per tree
was similar among the selected algarrobo forests (table 2).
Table
2. Stand structural characteristics of the
sampled sites.
Tabla 2. Características
estructurales de los sitios muestreados.
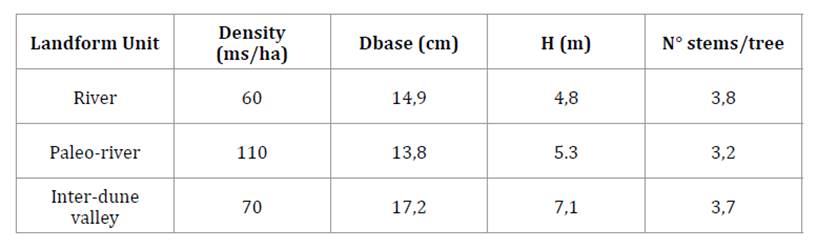
ms:
multi-stemmed tree. Dbase: mean multi-stemmed tree basal diameter; H: mean
multi-stemmed tree height.
ms:
árbol multi-fustal. Dbase: diámetro basal promedio del árbol multi-fustal. H:
altura promedio del árbol multi-fustal.
For each
multi-stemmed tree, two samples were extracted with a gas-powered drill
(TED_262R, Tanaka Kogyo Co. Ltd, Chiba, Japan), at approximately 50 cm above
the ground from the stem with the highest basal diameter. Samples were
air-drained, mounted on a wooden support, and polished with progressively finer
sandpaper to highlight wood anatomy. Rings were dated following the Schulman
criteria for the Southern Hemisphere tree species (25).
Tree-ring widths were measured from pith to bark with a 0.01 mm resolution Velmex
table connected to a computer. The correct dating of these measurements was
statistically validated with COFECHA software (13).
Then, we estimated individual tree diametric increments by averaging ring
measurements from samples belonging to each algarrobo tree and multiplying
these values by two. Finally, we calculated the cumulative diametric increment (CDI) as the sum of the current annual increments in diameter for each
biological year. This information allowed us
to estimate the time, in absolute tree age, required for the stem to reach the
minimum diameter corresponding to forest products in CMD algarrobo woodlands,
based on available information for the studied area (poles: stem ø= 10 cm) (19). When the core
center was not reached, tree age was estimated through a geometric method (10). We used ANOVA with
post-hoc Fisher’s least significant difference test through the Infostat software (9) to compare tree age
data across landform units. Five trees were excluded from this analysis since
they did not reach minimum CDI values of 10 cm (figure 2).
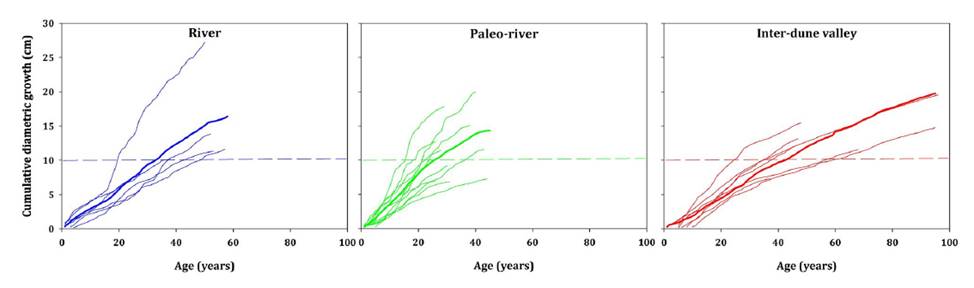
Dashed horizontal lines show the minimum stem ø = 10
cm. / Las líneas puntuadas indican el ø mínimo del fuste = 10 cm.
Figure 2. Cumulative
diameter increment for the River, Paleo-river, and Inter-dune valley landform
units.
Figura 2. Crecimiento
diamétrico cumulativo para las unidades de paisaje de Ripario, Paleo-ripario, y
valle inter-medanoso.
Biological
rotation age of the sampled algarrobo trees was estimated considering raw ring
widths converted to basal area increment (BAI). First, tree-ring measurements
from the two radii, corresponding to the two samples extracted for each tree,
were averaged at tree level to obtain individual ring-width chronologies. Then,
the AGE routine included in the DPL software
(Dendrochronological Program Library) (14) was
used to estimate current (CBAI) and mean (MBAI) basal area, according to the
following equation:
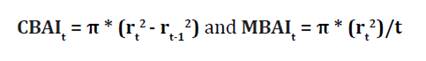
where:
rt represents ring
width in year t. Biological rotation age (cutting cycle period) was determined
at the age when MBAI and CBAI intersected (5).
Results
Tree-ring width
chronologies were constructed using 36 samples corresponding to 24 N.
flexuosa multi-stemmed trees. The relatively low number of selected trees
at River and Inter-dune valley depended on landowner sampling permission.
Multi-stemmed algarrobo site chronologies spanned from 44 (Paleo-river unit) to
96 years (Inter-dune valley unit). Mean annual radial growth rates oscillated
between 1.27 (Inter-dune valley unit) and 1.77 mm/yr (Paleo-river unit). Mean
correlation values (MC) between individual tree-ring chronologies ranked from
0.446 (Inter-dune valley unit) to 0.490 (Paleo-river unit), with all values
being significant at p < 0.05 (table 3).
Table
3. Characteristics of the tree-ring
chronologies.
Tabla 3. Características
de las cronologías de ancho de anillo.
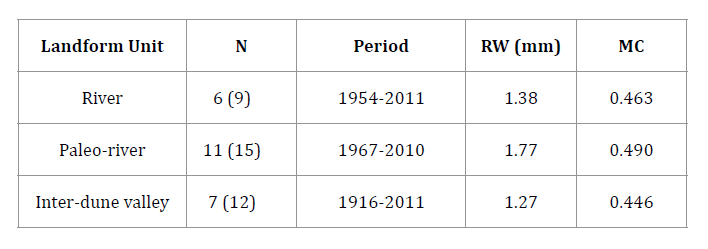
N = Number of sampled trees per site (in
parenthesis: total number of dendrochronological samples); Period = time range
of the sampled cores; RW = mean ring-width value; MC = mean correlation between
individual tree-ring series at each stand.
N = número de los árboles muestreados (entre
paréntesis: número total de las muestras dendrocronológicas); Period =
intervalo temporal de las muestras; RW = valor de ancho de anillo promedio; MC
= correlación promedio entre las series dendrocronológicas individuales para
cada sitio.
CDI analysis
revealed that multi-stemmed trees achieved, on average, a minimum diameter of
10 cm at different ages: 37 years for River unit, 24 years for Paleo-river
environment, and 44 years for Inter-dune valley landform (figure
2).
Statistically
significant differences emerged regarding tree age corresponding to CDI ø= 10
cm among Paleo-river units on one side, and River and Inter-dune valley
landforms on the other (one-way ANOVA, F = 5.40, p = 0.0161, df =
2, n = 19; figure 3).
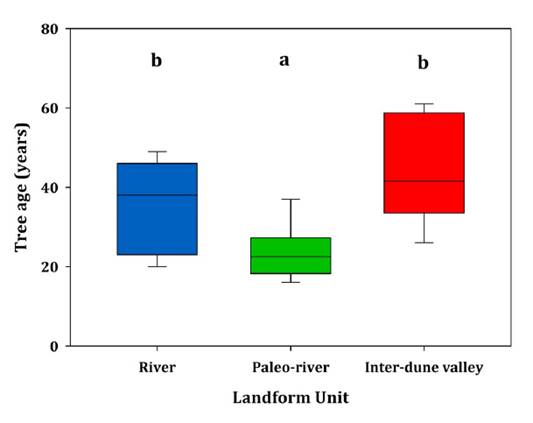
Each box shows the values within one interquartile
distance (ID 25% above and below the median). The median is shown as a black
horizontal bar in the boxes. Whiskers represent values reaching 1.5 times the
IDs. Different letters indicate significant differences at p < 0.05.
Cada caja muestra los valores comprendidos en la
distancia de un intercuartil (ID 25% por encima y por debajo del promedio). El
promedio se muestra como línea negra. Los bigotes corresponden a un valor de
1,5 veces la distancia de un ID y se muestran como líneas negras. Letras
diferentes indican diferencias significativas por p < 0,05.
Figure 3. Box
and whiskers plot for tree age corresponding to the minimum harvestable
diameter of 10 cm across landform units.
Figura 3. Gráficos
de caja-bigote para los valores del diámetro mínimo maderable de 10 cm según la
unidad de paisaje.
Landform
heterogeneity was reflected in biological rotation age among the selected
units. In this sense, the cutting cycle period occurred at ages 53 and 41 years
at River and Paleo-river units, corresponding to stem diameters of 19 and 18
cm, respectively. In contrast, at the Inter-dune valley landform, the
culmination age exceeded 96 years (stem ø = 24.5 cm), corresponding to the age
of the two oldest sampled trees (figure 4).
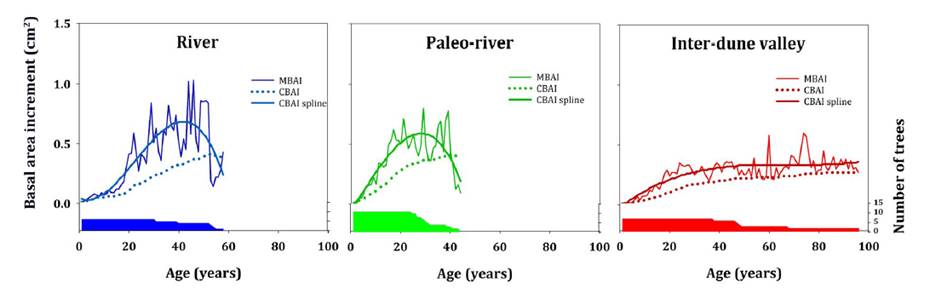
Coloured areas show the number of trees used for
estimating MBAI and CBAI.
Las áreas coloreadas indican el número de árboles
utilizados en el cálculo de MBAI y CBAI.
Figure 4. Mean
(MBAI) and current (CBAI) annual basal area increments in relation to tree ages
across landform units.
Figura 4. Incrementos
anuales en área basal promedio (MBAI) y corriente (CBAI) en relación a la edad
del árbol según la unidad de paisaje.
Discussion
In this study,
we employed dendrochronological methods to reconstruct growth dynamics of
multi-stemmed N. flexuosa across three distinct landform units within
the CMD. This research provides pioneering evidence of how topographical and
edaphic factors influence growth rates of multi-stemmed algarrobo trees in the
studied area. Our investigation delivered novel insights concerning the most
important forest resource that sustains local communities residing in arid
central-west Argentina. The low number of examined multi-stemmed N. flexuosa
individuals at two sites is not considered an analytic limitation, aimed to
explore for the first time tree growth form-dependent dynamics at regional
scale. Future studies increasing site repetitions as well total sampled tree
number could validate our findings.
Our analyses
demonstrated that landform variations were mirrored by growth rates of
multi-stemmed algarrobo trees, evidenced by differences in the time required to
attain harvestable stem diameters, and the length of the cutting cycle period.
Previous dendrochronological assessments suggested that soil attributes and
environmental heterogeneity play pivotal roles in shaping moisture availability
in the CMD districts, thereby influencing growth dynamics of algarrobo woodlands
(21). In this sense, it is worth
mentioning that N. flexuosa exhibits characteristics of a facultative
phreatophyte species, with distinctive root architecture that enables algarrobo
trees to access deep phreatic water while also exploring shallow soil horizons
(12). This trait allows N. flexuosa trees
to use both precipitation and groundwater for tree growth (15). Across the woodlands examined in our study,
the phreatic water table consistently lies at similar depths, approximately
between 4.5 and 6 meters (26), suggesting
that factors beyond water table accessibility, likely soil characteristics and
impact on water retention of superficial horizons, probably contributed to the
observed variations in tree growth. In arid ecosystems like the CMD, sandy
soils tend to facilitate deep water infiltration, whereas loamy edaphic layers
promote water retention in superficial horizons, resulting in enhanced stem
growth (17, 18, 21). Moreover, soil
permeability directly influences water storage capacity, with moderately
permeable soils exhibiting better precipitation retention compared to those
with higher permeability (18, 21). Both
of these edaphic attributes may collectively account for our research outcomes.
Previous
analyses have indicated that the biological rotation age of N. flexuosa is
growth-form dependent, with multi-stemmed trees reaching the cutting cycle
period at younger age than their one-stemmed counterparts (4). According to Alvarez et
al. (2011), multi-stemmed algarrobo trees could be harvested at
approximately 80 years of age. However, our study underscores that landform
clearly modulated the cutting cycle period. Our findings revealed different
values for biological rotation age across the examined stand. Regarding the
discrepancy observed for similar environments, that is, between the Inter-dune
valley stand analyzed in this research and the woodland examined by Alvarez et al. (2011), belonging to the same
landform unit, it could be hypothesized that variations in topographical
features, such as the presence (4) or
absence (our study) of a complete dune system surrounding the Inter-dune valley
site, may affect runoff processes and hence water availability. This landform
variability could potentially account for this particular divergent result,
although further investigations are warranted to explore the specific
topographic influence at a smaller spatial scale within the Inter-dune valley
unit, to address this difference more comprehensively.
At local scale,
our results indicated that wood extraction from multi-stemmed N. flexuosa in
CMD districts should consider landform heterogeneity. Specifically, while
multi-stemmed algarrobo trees in loamy Paleo-river soil and, to a lesser
extent, River units could be managed for pole production, pruning should be
avoided in Inter-dune valley N. flexuosa forest stands. This
recommendation is grounded in the exceptionally extended time required to reach
biological rotation age in this landform, which is approximately 100 years.
Additionally, the slow radial growth rates in the Inter dune valley environment
translated to an extended time window for obtaining forest products used by
local inhabitants in the studied area.
Conclusions
This research
sheds crucial light on a strategically vital forest resource for the
communities residing in the CMD area. These findings should be integrated with
the development of future forest management strategies for the semi-arid N.
flexuosa woodlands located in the central-western region of Argentina. On a
broader scale, our study underscores the paramount importance of considering
landform heterogeneity in the management of desert resprouting woody
vegetation.
Acknowledgments
The authors
warmly thank the Aguero and Cordoba families for allowing sampling in their
respective areas. Special thanks are due to Hugo Debandi and Alberto Ripalta
for field assistance. We thank the Direccion de Recursos Naturales Renovables
of Mendoza Province for allowing sampling.
1. Abraham, E.
M.; Prieto, M. R. 1999. Vitivinicultura y desertificación en Mendoza. En:
García Martínez, B. (Ed.), Estudios de historia y ambiente en América:
Argentina, Bolivia, México, Paraguay. IPGH-Colegio de México, México. 109-135.
2. Abraham, E.;
del Valle, H. F.; Roig, F.; Torres, L.; Ares, J. O.; Coronato, F.; Godagnone,
R. 2009. Overview of the geography of the Monte Desert biome (Argentina).
Journal of Arid Environments. 73(2): 144-153.
3. Alvarez, J.
A.; Villagra, P. E.; Cony, M. A.; Cesca, E.; Boninsegna, J. A. 2006. Estructura
y estado de conservación de los bosques de Prosopis flexuosa DC
(Fabaceae, subfamilia: Mimosoideae) en el noreste de Mendoza (Argentina).
Revista Chilena de Historia Natural. 79(1): 75-87.
4. Alvarez, J.
A.; Villagra, P. E.; Villalba, R.; Cony, M. A.; Alberto, M. 2011b. Wood
productivity of Prosopis flexuosa DC woodlands in the central Monte:
Influence of population structure and treegrowth habit. Journal of Arid
Environments. 75(1): 7-13.
5. Assman, E.
1970. The principles of forest yield study. Section B: Tree growth and form,
Pergamon Press. New York. 51-53.
6. Baldassini,
P.; Bagnato, C. E.; Paruelo, J. M. 2020. How may deforestation rates and
political instruments affect land use patterns and Carbon emissions in the
semi-arid Chaco, Argentina?. Land Use Policy. 99:
104985.
7. Bastin, J.
F.; Berrahmouni, N.; Grainger, A.; Maniatis, D.; Mollicone, D.; Moore, R.,
Patriarca, C.; Picard, N.; Sparrow, B.; Abraham, E. M.; Aloui, K.; Atesoglu,
A.; Attore, F.; Bassüllü, C.; Bey, A.; Garzuglia, M.; García-Montero, L. G.;
Groot, N.; Guerin, G.; Laestadius, L.; Lowe, A.; Mamane, B.; Marchi, G.;
Patterson, P.; Rezende, M.; Ricci, S.; Salcedo, I.; Sanchez-Diaz Paus, A.;
Stolle, F.; Surappaeva, V.; Castro, R. 2017. The extent of forest in dryland
biomes. Science. 356(6338): 635-638.
8. Bond, W. J.;
Midgley, J. J. 2001. Ecology of sprouting in woody plants: the persistence
niche. Trends in ecology & evolution. 16(1): 45-51.
9. Di Rienzo, J.
A.; Casanoves, F.; Balzarini, M. G.; González, L.; Tablada, M.; Robledo, Y. C.
2011. InfoStat versión 2011. Grupo InfoStat, FCA, Universidad Nacional de
Córdoba, Argentina. http://www. infostat.com.ar. 8: 195-199.
10. Duncan, R.
P. 1989. An evaluation of errors in tree age estimates based on increment cores
in kahikatea (Dacrycarpus dacrydioides). New Zealand Natural Sci. 16:
31-37.
11. FAO (Food
and Agriculture Organization). 1977. Carte mondiale de la désertification a
l’échelle de 1:25,000,000, Conférence des Nations
Unies sur la désertification, 29 août–9 septembre, A/CONF. 74/2.
12. Guevara, A.;
Giordano, C. V.; Aranibar, J.; Quiroga, M.; Villagra, P. E. 2010. Phenotypic
plasticity of the coarse root system of Prosopis flexuosa, a
phreatophyte tree, in the Monte Desert (Argentina). Plant and soil. 330(1-2):
447-464.
13. Holmes R. L.
1983. Computer-assisted quality control in tree-ring dating and measurement.
Tree-Ring Bulletin. 43: 69-78.
14. Holmes, R.
L. 1999. Dendrochronological Program Library (DPL). User’s Manual, Laboratory
of Tree-Ring Research, University of Arizona. Tucson. Arizona. USA.
15. Jobbágy, E.
G.; Nosetto, M. D.; Villagra, P. E.; Jackson, R. B. 2011. Water subsidies from
mountains to deserts: their role in sustaining groundwater-fed oases in a sandy
landscape. Ecological Applications. 21(3): 678-694.
16. Maestre, F.
T.; Benito, B. M.; Berdugo, M.; Concostrina-Zubiri, L.; Delgado-Baquerizo, M.;
Eldridge, D. J.; Guirado, E.; Gross, N.; Kéfi, S.; Le BagoussePinguet, Y.;
Ochoa-Hueso, R.; Soliveres, S. 2021. Biogeography of global drylands. New
Phytologist. 231(2): 540-558.
17. Massa, A.;
Quagliariello, G.; Martinengo, N.; Calderón, A.; Pérez, S. 2023. Growth and
slenderness index in sweet algarrobo, Neltuma flexuosa, according to the
vermicompost percentage in the substrate and seed origin. Revista de la
Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza.
Argentina. 55(2): 12-19. DOI: https://doi.org/10.48162/rev.39.105
18. Noy-Meir, I.
1973. Desert ecosystems: environment and producers. Annual review of ecology
and systematics. 4: 25-51.
19. Ongay
Ugarteche, O.; Marambio Fermani. S.; Corominas Day, M.; Lagos Slinik, S.
Acordinaro N. 2011. Manual de Bosques Nativos. Un aporte a la Conservación
desde la Educación Ambiental. Dirección de Recursos Naturales Renovables
Secretaría de Medio Ambiente Gobierno de Mendoza. 121 p.
20. Petrie, M.
D.; Bradford, J. B.; Hubbard, R. M.; Lauenroth, W. K.; Andrews, C. M.;
Schlaepfer, D. R. 2017. Climate change may restrict dryland forest regeneration
in the 21st century. Ecology. 98(6): 1548-1559.
21. Piraino, S.;
Abraham, E. M.; Diblasi, A.; Roig Juñent, F. A. 2015. Geomorphological-related heterogeneity
as reflected in tree growth and its relationships with climate of Monte Desert Prosopis
flexuosa DC woodlands. Trees. 29: 903-916.
22. Piraino, S.;
Abraham, E. M.; Hadad, M. A.; Patón, D.; Roig-Juñent, F. A. 2017. Anthropogenic
disturbance impact on the stem growth of Prosopis flexuosa DC forests in
the Monte desert of Argentina: A dendroecological approach. Dendrochronologia.
42: 63-72.
23. Rubio, M.
C.; Soria, D.; Salomón, M. A.; Abraham, E. 2009. Delimitación de unidades
geomorfológicas mediante la aplicación de técnicas de procesamiento digital de
imágenes y SIG. Área no irrigada del departamento de Lavalle, Mendoza.
Proyección. 2: 1-33.
24. Schild, J.
E.; Vermaat, J. E.; van Bodegom, P. M. 2018. Differential effects of valuation
method and ecosystem type on the monetary valuation of dryland ecosystem
services: A quantitative analysis. Journal of arid environments. 159: 11-21.
25. Schulman, E.
1956. Dendroclimatic changes in semiarid America. University of Arizona Press, Tucson.
26. Soria, D.;
Rubio, C.; Abraham, E. M. 2011. Sitio piloto en la Región Centro Oeste. En:
“Evaluación de la desertificación en Argentina. Resultados del proyecto
LADA/FAO”. FAO, PAN, UNEP, GEF, LADA y SAyDS. Gráfica Latina. Buenos Aires,
Argentina. 205-246.
27. Vázquez, D.
P.; Alvarez, J. A.; Debandi, G.; Aranibar, J. N.; Villagra, P. E. 2011.
Ecological consequences of dead wood extraction in an arid ecosystem. Basic and
Applied Ecology. 12(8): 722-732.
28. Villagra, P.
E.; Defossé, G. E.; Del Valle, H. F.; Tabeni, S.; Rostagno, M.; Cesca, E.;
Abraham, E. 2009. Land use and disturbance effects on the dynamics of natural
ecosystems of the Monte Desert: Implications for their management. Journal of
Arid Environments. 73(2): 202-211.
29. Wang, L.;
Jiao, W.; MacBean, N.; Rulli, M. C.; Manzoni, S.; Vico, G.; D’Odorico, P. 2022.
Dryland productivity under a changing climate. Nature Climate Change. 12(11):
981-994.
30. Ware, I. M.;
Ostertag, R.; Cordell, S.; Giardina, C. P.; Sack, L., Medeiros, C. D.; Inman,
F.; Litton, C. M.; Giambelluca, T.; John, G. P.; Scoffoni, C. 2022.
Multi-stemmed habit in tres contributes climate resilience in tropical dry
forest. Sustainability. 14(11). 6779.